Ein Nervensignal ist ein elektrisches Signal und wird
durch den Strom von Ionen (in H20 gelöste Salze) durch
Kanäle in der Membran hervorgerufen. Solche Kanäle, auch Kanalproteine
genannt, sind von Bedeutung für die Weiterleitung von Informationen,
und sie sind nicht für alle Ionen gleich durchlässig. Wäre
die Membran für alle gleich durchlässig so würde sie sich
im chemisch-elektrischen Gleichgewicht befinden. Das heißt, die Konzentration
von positiven Natrium- und Kaliumionen und negativen Chlorionen im Innern
und Äußeren der Membran wäre gleich.
Dies ist nicht so, da Kaliumionen stärker durchgelassen
werden als Natriumionen. Chlorionen können die Membran fast überhaupt
nicht durchqueren. Daher herrscht im Ruhezustand der Membran ein Konzentrationsunterschied
der drei wichtigsten Ionenarten: Natrium, Kalium und Chlor. Die Flüssigkeit
außerhalb der Nervenelle hat eine zwölf mal höhere Natriumkonzentration
als innen. Innerhalb ist die Konzentration an Kaliumionen vierzig mal höher
als außen.
Durch die stärkere Durchlässigkeit der Membran für Kaliumionen können diese in höherer Zahl ausströmen als negative Ionen. Dadurch entsteht ein Ungleichgewicht an positiven und negativen Ladungen zwischen dem Inneren und dem Äußeren der Zelle, was ein elektrisches Feld aufbaut. Dieses kann, wenn der Ladungsunterschied eine bestimmte Größe erreicht hat, den Diffusionsstrom der Ionen kompensieren, und es tritt ein Ruhezustand ein. Im Ruhezustand ist eine Potentialdifferenz von ungefähr -70 mV vom Inneren gegenüber dem Äußeren gegeben. Trotz der teilweise durchlässigen Membran wird das Konzentrationsgefälle zwischen innen und außen durch spezielle Pumpenproteine (spezielle Moleküle) aufrechterhalten, die Ionen auf die andere Seite transportieren. Eine derartige Pumpe ist die Natriumpumpe.
Siehe Abb.3.2: Fortpflanzung
des Nervensignals entlang eines unmyelinisierten Axons.
Befindet sich ein Axon im Ruhezustand, herrscht also ein
Potentialunterschied von -70 mV vom Inneren gegenüber dem Äußeren.
Kommt dann ein elektrisches Nervensignal zur Zelle, so bewirkt sein elektrisches
Feld, dass sich Natriumkanäle öffnen und somit Natriumionen von
außen nach innen, aber keine Kaliumionen nach außen strömen
können. Hierdurch verringert sich das negative Membranenpotential,
was die Ursache dafür ist, dass sich noch weitere Natriumkanäle
öffnen und ein relativ hoher Strom von Natriumionen in die Zelle gelangt,
so dass sich das Membranenpotential weiter verringert, positiv wird und
bis auf einen Wert von 30 mV ansteigt.
Jetzt werden Kaliumkanäle geöffnet, Kaliumionen
gelangen nach außen, so dass der Ruhezustand wieder hergestellt werden
kann. Die Phase, in der das Zellpotential unter dem Ruhepotential liegt,
nennt man Refraktärphase. Zu dieser Zeit reagiert die Zellmembran
nur schwer auf Reize. Die Ionenpumpen sind jetzt voll ausgelastet damit,
die über die Kanäle ein- bzw. ausgeströmten Ionen wieder
nach innen bzw. nach außen zu bringen.
Wichtig für das Verständnis der Weiterleitung
des Nervensignals ist, dass das Aktionspoten-tial überall die gleiche
Form und Amplitude besitzt. Daher kann kein Einzelsignal eine Information
tragen. Die Nachricht über die Stärke des Ausgangsreizes wird
durch die Frequenz des Aktionspotentials codiert. Durch diese Frequenzcodierung
kann das Aktionspotential über lange Strecken unverändert übertragen
werden.
Die Fortbewegungsgeschwindigkeit eines Nervensignals in
einem unmyelinisierten Axon hängt vom Durchmesser des Querschnitts
des Axons ab. Die Nervenfaser muß, um Signale doppelt so schnell
leiten zu können, die vierfache Dicke haben. Myelinisierte Nervenfasern
benötigen bei gleicher Informationsübertragungsgeschwindigkeit
weniger Volumen. Ihre Dicke ist proportional zur Übertragungsgeschwindigkeit.
Das menschliche Rückenmark müßte mehrere Meter dick sein,
wäre es unmyelinisiert.
Eine solche Myelinschicht umgibt die Axone jedoch nicht
vollständig, sondern hat in ein-Meter-Abständen Einschnürungen,
die Ranvier-Schnürringe, an denen die Schicht fehlt. Bei unmyelinisierten
Zellen ergibt sich durch Einstrom von Natriumionen eine Polarisation und
darauf durch Ausstrom von Kaliumionen wieder eine Depolarisation des jeweiligen
Abschnitts der Nervenfaser.
Die elektrischen Feldlinien eines Nervensignals sind
um so dichter, je näher der noch unpolarisierte Abschnitt des Axons
am bereits depolarisierten Bereich liegt. Deshalb bewirkt das elektrische
Feld die Öffnung von Natriumkanälen in der Nähe des depolarisierten
Abschnitts und ermöglicht hierdurch die Weiterleitung des Aktionspotentials.
Die Feldlinien können die isolierende Myelinhülle nicht durchdringen,
sondern springen von einem depolarisierten Abschnitt zum nächsten
und polarisieren dort die Membran durch Öffnung von Natriumkanälen.
So gelangt dann das Aktionspotential viel schneller von
einem Schnürring zum nächsten. Die Gehirne von Säugetieren
könnten ohne Myelin nicht so kompakt aufgebaut sein.
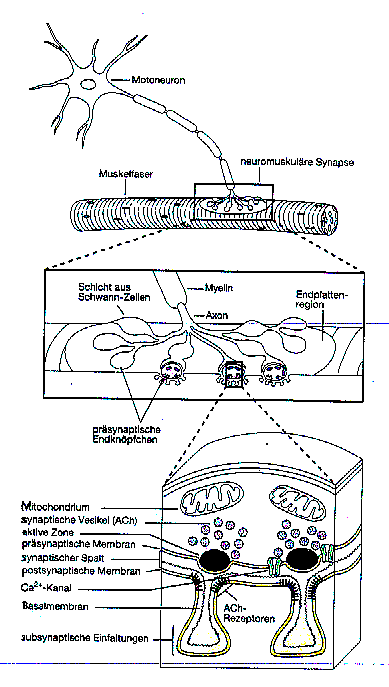
Diese Vorgänge finden alle durch Signalleitung über
eine erregende Synapse statt. Eine andere Art von Synapsen sind hemmende
Synapsen. Sie bewirken den umgekehrten Fall. Es wird eine Erniedrigung
des negativen Ruhepotentials ausgelöst und damit die Auslösung
von Aktionspotentialen an der Basis des Neurons verhindert.
Die Stärke der Verbindung durch eine Synapse ist
veränderbar. Werden über eine Synapse sehr oft Informationen
weitergeleitet, ist ihre Effektivität ausgeprägter als die einer
weniger aktiven Synapse, das heißt, Synapsen sind lernfähig.
Wahrscheinlich ist dies der bedeutendste Vorgang des Lernens der Lebewesen
mit Neuralsystem. Bei den meisten künstlichen neuronalen Netzen dienen
diese Eigenschaften als entscheidende Grundlage.
| Proseminar-Startseite |
{kind=link}
{kind=link}
{kind=link}
{kind=link}